- Biology Article
- Griffith Experiment Genetic Material

Griffith Experiment and Search of Genetic Material

The search for Genetic material started during the mid-nineteenth century. The principle of inheritance was discovered by Mendel. Based on his investigation, Mendel concluded that some ‘factors’ are transferred from one generation to another. Mendel’s Law of Inheritance was the basis for the researchers on genetic material. Keeping his conclusions in mind, scientists who came after him, focused on chromosomes in search of genetic material. Even though the chromosomal components were identified, the material which is responsible for inheritance remained unanswered. It took a long time for the acceptance of DNA as the genetic transformation. Let’s go through a brief account of the discovery of genetic material and Griffith experiment.
Griffith Experiment & Transforming Principle
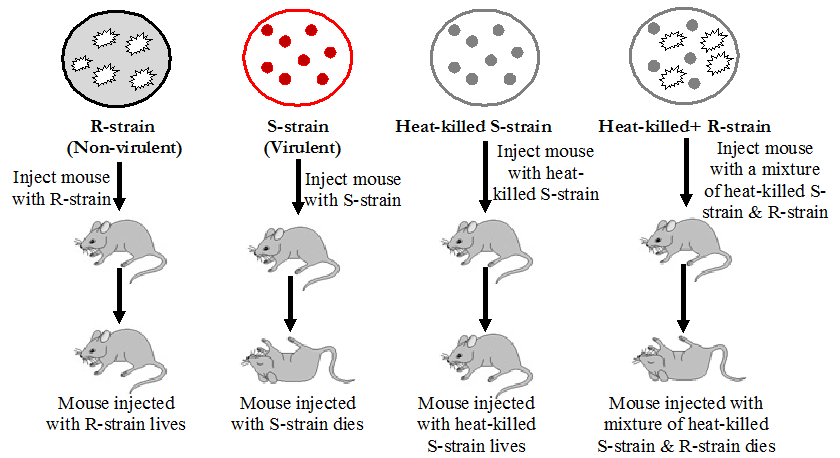
Griffith experiment was a stepping stone for the discovery of genetic material. Frederick Griffith experiments were conducted with Streptococcus pneumoniae.
During the experiment, Griffith cultured Streptococcus pneumoniae bacteria which showed two patterns of growth. One culture plate consisted of smooth shiny colonies (S) while other consisted of rough colonies (R). The difference was due to the presence of mucous coat in S strain bacteria, whereas the R strain bacteria lacked them.
Experiment: Griffith injected both S and R strains to mice. The one which was infected with the S strain developed pneumonia and died while that infected with the R strain stayed alive.
In the second stage, Griffith heat-killed the S strain bacteria and injected into mice, but the mice stayed alive. Then, he mixed the heat-killed S and live R strains. This mixture was injected into mice and they died. In addition, he found living S strain bacteria in dead mice.

Conclusion: Based on the observation, Griffith concluded that R strain bacteria had been transformed by S strain bacteria. The R strain inherited some ‘transforming principle’ from the heat-killed S strain bacteria which made them virulent. And he assumed this transforming principle as genetic material.
DNA as Genetic Material
Griffith experiment was a turning point towards the discovery of hereditary material. However, it failed to explain the biochemistry of genetic material. Hence, a group of scientists, Oswald Avery, Colin MacLeod and Maclyn McCarty continued the Griffith experiment in search of biochemical nature of the hereditary material. Their discovery revised the concept of protein as genetic material to DNA as genetic material .
Avery and his team extracted and purified proteins, DNA, RNA and other biomolecules from the heat-killed S strain bacteria. They discovered that DNA is the genetic material and it is alone responsible for the transformation of the R strain bacteria. They observed that protein-digesting enzymes (proteases) and RNA-digesting enzymes (RNases) didn’t inhibit transformation but DNase did. Although it was not accepted by all, they concluded DNA as genetic material.
Frequently Asked Questions on Griffith Experiment
What was griffith’s experiment and why was it important.
Griffith’s experiment was the first experiment which suggested that bacteria can transfer genetic information through a process called transformation.
What is the conclusion of Griffith experiment?
The experiment concluded that bacteria are capable of transfering genetic information through transformation.
What was the most significant conclusion of Griffith’s experiments with pneumonia in mice?
The experiment conducted by Griffith found that bacteria are capable of transfering genetic information through transformation.
What did Frederick Griffith want to learn about bacteria?
Frederick Griffith wanted to learn if bacterial transformation was possible.
How did the two types of bacteria used by Griffith differ?
Griffith used two strains of pneumococcus (Streptococcus pneumoniae) bacteria: a type III-S and a type II-R.

Put your understanding of this concept to test by answering a few MCQs. Click ‘Start Quiz’ to begin!
Select the correct answer and click on the “Finish” button Check your score and answers at the end of the quiz
Visit BYJU’S for all Biology related queries and study materials
Your result is as below
Request OTP on Voice Call
Leave a Comment Cancel reply
Your Mobile number and Email id will not be published. Required fields are marked *
Post My Comment
Hi! This is well explained. I love it
Register with BYJU'S & Download Free PDFs
Register with byju's & watch live videos.
Talk to our experts
1800-120-456-456
Griffith Experiment

Griffith Experiment: An Introduction
It may come as a surprise that less than a century ago, even the most educated members of the scientific community were unaware that DNA was a hereditary material. Frederick Griffith conducted a series of experiments with Streptococcus pneumonia bacteria and mice in 1928 and concluded that the R-strain bacteria must have picked up a "transforming principle" from the heat-killed S bacteria, allowing them to "transform" into smooth-coated bacteria and become virulent.
In this article, we'll look at one of the classic experiments that led to the discovery of DNA as a genetic information carrier.
Who was Frederick Griffith?
The "Griffith's Experiment," carried out by English bacteriologist Frederick Griffith in 1928, described the transformation of a non-pathogenic pneumococcal bacteria into a virulent strain.
Griffith combined living non-virulent bacteria with a heat-inactivated virulent form in this experiment.
He was the first to discover the "transforming principle," which led to the discovery of DNA as a carrier of genetic information.
He suggested that bacteria can transfer genetic information via a process known as transformation.
Griffith's goal was not to identify the genetic material but to create a vaccine against pneumonia. In his experiments, Griffith used two related strains of bacteria known as R and S.
Griffith's work was expanded by Avery, MacLeod, and McCarty.
R Strain And S Strain Bacteria
Streptococcus pneumonia comes in several types or strains. Griffith chose two different strains for his experiment.
One strain of bacteria has smooth surfaces and is known as the smooth strain (S strain), while the other has rough surfaces and is known as the rough strain (R strain).
Bacteria of the S strain have smooth surfaces because they produce a polysaccharide protective coating that forms the outermost layer.
Apart from the morphological differences, Griffith discovered another significant difference between the S and R strains of bacteria, i.e., the S strain is the "virulent" strain capable of causing death in mice, whereas the R strain is the "nonvirulent" strain that will not cause death in mice.
Griffith observed that when he injected these bacteria into mice, the mice infected with the virulent S strain died from pneumonia, whereas the mice infected with the nonvirulent R strain survived.

R Strain and S Strain of Streptococcus Pneumonia
Griffith’s Transformation Experiment
Griffith was researching the possibility of developing a pneumonia vaccine.
He used two strains of pneumococcus (Streptococcus pneumonia) bacteria that infect mice – a virulent (causing disease) S (smooth) strain and a non-virulent type R (rough) strain.
The S strain produced a polysaccharide capsule that protected itself from the host's immune system, resulting in the host's death, whereas the R strain lacked that protective capsule and was defeated by the host's immune system.
Griffith attempted to inject mice with heat-killed S bacteria as a part of his research (i.e., S bacteria that had been heated to high temperatures, causing the cells to die). The heat-killed S bacteria, but unsurprisingly, did not cause disease in the mouse.
When harmless R bacteria were combined with harmless heat-killed S bacteria and injected into a mouse, the experiments took an unexpected turn.
Not only did the mouse develop pneumonia and die, but Griffith discovered living S bacteria in a blood sample taken from the dead mouse.
He concluded that some factor or biomolecule from the heat-killed S bacteria had entered the living R bacteria, allowing them to synthesise a polysaccharide coating and become virulent. As a result, this factor "transformed" the R bacteria into S bacteria.
Griffith called this factor the "transforming principle," concluding that it carried some genetic material from the S bacteria to the R bacteria.
This process is now known as bacterial transformation and is used in a variety of significant genetic engineering applications.

Griffith Experiment Diagram
Impact of The Griffith Experiment
One of the characteristics of hereditary material is a changing phenotype . Griffith referred to the phenotypic-changing factor as the transforming principle.
His work on the transforming principle received the most attention, but only after a group of Canadian and American scientists set out to investigate the chemical nature of the transforming principle in Oswald Avery's laboratory.
Avery's group concluded in their studies that deoxyribonucleic acid was the molecule identified by Griffith as the transforming principle after conducting numerous experiments.
The implications of this discovery are farfetched because it was made at a time when scientists considered protein molecules to be genetic material.
DNA, or deoxyribonucleic acid, is now recognised as the molecule that encodes all cell functions and transmits genetic information from parent to offspring in almost every living species .
In the 1940s, however, DNA was thought to be a less qualified candidate for genetic material. Avery and colleagues' research on Griffith's experiment provided the first solid evidence that DNA could be the genetic material.
Griffith's ultimate goal was to find a way to cure pneumonia. Griffith inoculated mice with various strains of pneumococci to see if they would infect and eventually kill the mice. Griffith concluded that heat-killed virulent bacteria transformed living, non-virulent bacteria into virulent bacteria. He performed his experiment on the two strains of Streptococcus pneumonia, which differ from each other due to the presence of a polysaccharide coat.
Griffith's findings were published in the Journal of Hygiene. In 1928, his experiments with mice led to his major discovery of bacterial transformation. Griffith's experiment discovered that bacteria can transfer genetic information through transformation.
FAQs on Griffith Experiment
1. Explain the Oswald Avery Experiment.
Avery and his colleagues conducted additional research on the virulent S strain of Streptococcus pneumonia. They were aware that the potential carriers of genetic material were proteins, RNA, or DNA. When the mixtures were treated with protein-digesting or RNA-digesting enzymes, the DNA remained intact and was capable of transforming R bacteria into S bacteria. However, when the DNA in these mixtures was broken down with DNase, the genetic material could not be passed from the heat-killed S bacteria to the live R bacteria, preventing transformation. As a result, Avery and his colleagues concluded that the transforming principle described by Griffith had to be DNA.
2. What are the characteristics of genetic material?
Any substance that forms the genetic material must fulfil some essential requirements:
It must be stable.
It should be able to carry and transcribe information which is required to control the processes.
It should be able to replicate itself and remain unchanged while passing down from one generation to another.
It must be able to mutate itself to provide variations.
A genetic material must be able to store the information, transmit it, replicate it and provide variation.
DNA fulfils all the above-mentioned requirements and hence acts as genetic material.
3. Define Horizontal Gene Transfer.
Horizontal gene transfer (HGT) is the exchange of genetic information between organisms, which includes the spread of antibiotic resistance genes among bacteria (except those passed down from parent to offspring), thereby, fueling pathogen evolution.
Bacterial horizontal gene transfer occurs via three mechanisms: transformation, transduction, and conjugation. Conjugation is the primary mechanism for the spread of antibiotic resistance in bacteria, and it is critical in the evolution of the bacteria that degrade novel compounds such as pesticides created by humans, as well as in the evolution, maintenance, and transmission of virulence.
Lessons and Courses on Microbiology
Bacterial Transformation (The Transforming Principle)
Fredrick Griffith showed in 1928 that deoxyribonucleic acid (DNA) is the genetic material of the cell. His work was unique because it was the first evidence that showed that DNA is the genetic material of the cell. Other scientists that also carried out experiments to establish that DNA is the genetic material include Oswald Avery , Martha Chase and Alfred Hershey amongst others. Frederick Griffith was able to transform apathogenic bacteria (i.e. harmless bacteria) to virulent pathogenic bacteria in his transformation experiment.
Gregor Mendel’s work on hereditary to determine the mechanism of genetic inheritance set the stage for the development of the field known as genetics. And scientists over the years have built on these earlier experiments to unravel the genetic code of living organisms as well as manipulate same for other beneficial and economic purposes.
Friedrich Miescher was the first to describe the DNA in 1869, long before Griffith discovered DNA as the main genetic material of the cell. DNA is an integral part of the cell and it is exclusively found in the chromosomes where it is wrapped with histone proteins. The DNA or gene as it is often called encodes gene sequences that coordinate the expression of specific protein molecules in the cell; and they are passed on from parents to their offspring during reproduction.
Frederick Griffith carried out his experiment with mice and Streptococcus pneumoniae (pneumococcus bacteria) , the causative agent of pneumonia in humans. And he showed that there was a transforming principle in bacterial genome. Griffith called this transforming principle deoxyribonucleic acid (DNA) . He postulated that genetic information could somehow be transferred between different strains of bacteria (particularly amongst pathogenic and non-pathogenic organisms) in a systematic manner; and this could make a non-pathogenic organism to become pathogenic. S. pneumoniae (pneumococcus) occur in two different forms: as virulent strain and as non-virulent strain.
Griffith called the virulent pneumococcus strain S-strain and the non-virulent strain R-strain . The S-strain (virulent strains) of pneumococcus has a smooth polysaccharide capsule that play critical role in the pathogenicity and/or virulence of the bacterium while the R-strain (non-virulent) lacks the polysaccharide capsule. The R-strain has a rough appearance unlike the virulent strain because it lacks the polysaccharide capsule which is present in the later (i.e. the S-strain).
Griffith injected cultures of his rough strain (R-strain) and smooth strain (S-strain) into different groups of mice and allowed them for some days for pathogenicity to take place. He discovered that the mouse injected with the R-strain survived the inoculation while the mouse injected with the S-strain died after a few days ( Figure 1 ). Griffith went on to determine whether the viability of the smooth pneumococcus strain was required for the pathogenicity of the virulent strain.
He wanted to discern whether the factor present in the living smooth pneumococcus bacteria that causes disease (pneumonia) could be transferred to the nonpathogenic rough pneumococcus. To do this, Griffith boiled or heated the virulent S-strain for a short period of time in order to kill the virulent pneumococcus bacteria.
Griffith reasoned whether or not the heated (killed) virulent S. pneumoniae strain could cause disease in the killed state. He injected the heat killed S-strain into the mouse and allowed it for some days. On close observation, Griffith discovered that the mouse survived the inoculation of the heat-killed, smooth pneumococcus strain. Neither the rough pneumococcus bacteria nor the heat-killed smooth pneumococcus bacteria were capable of causing disease on their own; and this could be because they both lack the disease-causing capability which is present only in the smooth pneumococcus bacteria (i.e. the live S-strain).
In another experiment, Griffith inoculated mice with a mixture of the heat-killed virulent pneumococcus bacteria and lives but non-virulent R-strain; and he discovered that the mouse injected with the mixture of heat-killed pneumococcus bacteria and live but non-virulent pneumococcus bacteria survived. Heat denatures protein; and since the mice inoculated with heated-killed S-strain survived the injection, Griffith reasoned that some proteins within the chromosome of the live S-strain could be responsible for causing pneumonia in the mice. Frederick Griffith hypothesized that the live but non-virulent R-strain bacteria had by some means been transformed by the heat-killed S-strain smooth bacteria; and he postulated that some factors may be responsible for transforming the R-strain bacteria into virulent pneumococcus bacteria.
Griffith called the factor responsible for the disease-causing capability in the organism transforming principle , and the process of transformation he called genetic transformation. When pneumococcus bacteria were recovered from the dead mice, Griffith cultured them and found living, smooth pneumococcus bacteria growing on the media. He reasoned that the only way for this to have occurred was if the rough living pneumococcus bacteria (i.e. the R-strain) were transformed to become smooth virulent pneumococcus bacteria. The heating of the S-strain causes rupture on the cell wall of the organism which caused the organism to release its cellular constituents; and some of which were absorbed by the non-pathogenic strain that led to their transformation.
Once inside the cell, the cellular components of the heat-killed bacteria then transformed the living rough bacteria cells into living, smooth bacteria. The exact molecule(s) that make up the transforming factor were not known as at this time. This evidence pointed to DNA as the genetic material even though little was known about the nucleic acids (particularly DNA) as at the time of Griffiths experiment. However, in 1944, Oswald T. Avery and other notable scientists built on the experiments of Frederick Griffith and concluded that the transforming principle responsible for Griffith’s results was deoxyribonucleic acid ( Table 1 ).
Table 1. Summary of the discovery of DNA as the genetic material
While Frederick Griffith used bacteria (particularly the smooth and rough strains of pneumococcus) to establish the principle of bacterial transformation ; the later scientists including Avery Oswald , Alfred Hershey and Martha Chase used viruses i.e. bacteriophages to confirm Griffith’s transformation principle and establish that the genetic material of the cell is deoxyribonucleic acid (DNA).
The work of Frederick Griffith , Avery Oswald and Martha Chase and Alfred Hershey amongst other notable scientists established that DNA is the generic material and an integral part of the genes and chromosomes of living system; and their discoveries laid to rest the earlier claims that the genetic information or material of cells are contained in the cell proteins. Fredrick Griffith’s discovery of DNA as the genetic material was bacteriologically-based, but those of Avery Oswald and colleagues were based on biochemical and structural studies.
Alberts B , Bray D , Johnson A , Lewis J , Raff M , Roberts K and Walter P (1998). Essential Cell Biology: An Introduction to the Molecular Biology of the Cell. Third edition. Garland Publishing Inc., New York.
Ausubel, F.M., Brent, R., Kingston, R.E., Moore, D.D., Seidman, J.G., Smith, J.A., Struhl, K., eds (2002). Short Protocols in Molecular Biology, 5th edn. John Wiley & Sons, New York.
Bains W (1998). Biotechnology: From A to Z. 2nd ed. Oxford University Press, New York, USA.
Branden C and Tooze J (1998). Introduction to protein structure. A non-technical introduction to protein structure. New York: Garland Press.
Chen I and Dubnau D (2004). DNA uptake during bacterial transformation. Nat. Rev. Microbiol. 2 (3): 241–249.
Cooper G.M and Hausman R.E (2004). The cell: A Molecular Approach. Third edition. ASM Press.
Dale J (2003). Molecular genetics of bacteria. Jeremy W. Dale and Simon Park (4 th eds.). John Wiley & Sons Ltd, West Sussex, UK. Pp. 312-313.
Dale J (2003). Molecular genetics of bacteria. Jeremy W. Dale and Simon Park (4 th eds.). John Wiley & Sons Ltd, West Sussex, UK. Pp.
Glick B.R and Pasternak J.J (2003). Molecular Biotechnology: Principles and Applications of Recombinant DNA. ASM Press, Washington DC, USA.
Hames B.D and Rickwood D (1998). Gel Electrophoresis of Proteins: A Practical Approach 3 rd Edition. The Practical Approach Series, Oxford University Press
Latha C.D.S and Rao D.B (2007). Microbial Biotechnology. First edition. Discovery Publishing House (DPH), Darya Ganj, New Delhi, India.
Lewis R (2007). Human Genetics: Concepts and Applications. Seventh edition. McGraw-Hill Companies, Inc, New York, USA.
Lodish H, Berk A, Matsudaira P, Kaiser C.A, Kreiger M, Scott M.P, Zipursky S.L and Darnell J (2004). Molecular Cell Biology. Fifth edition. Scientific American Books, Freeman, New York, USA.
Madigan M.T., Martinko J.M., Dunlap P.V and Clark D.P (2009). Brock Biology of Microorganisms, 12 th edition. Pearson Benjamin Cummings Inc, USA.
Tamarin Robert H (2002). Principles of Genetics. Seventh edition. Tata McGraw-Hill Publishing Co Ltd, Delhi.
Thieman W.J, Palladamo M.A and Thieman W (2003). Introduction to Biotechnology. Benjamin Cummings, San Francisco, CA.
Share this:
Discover more from #1 microbiology resource hub.
Subscribe to get the latest posts to your email.
Type your email…
Related Posts

Introduction to Biotechnology

GENERAL SIGNIFICANCE / IMPORTANCE OF FUNGI
Leave a reply cancel reply.
Subscribe now to keep reading and get access to the full archive.
Continue reading
This page has been archived and is no longer updated
Discovery of DNA as the Hereditary Material using Streptococcus pneumoniae
No one could have predicted that experiments designed to understand bacterial pneumonia would lead to the discovery of DNA as the hereditary material. In the early part of the twentieth century, before the advent of antibiotics, pneumococcal infections claimed many more lives than they do today. Researchers on both sides of the Atlantic were thus actively engaged in studying Streptococcus pneumoniae , the bacterium responsible for clinical infections. Early on, it became apparent that multiple strains of S. pneumoniae were responsible for causing bacterial pnuemonia. Researchers also noted that patients developed antibodies to the particular strain, or serotype, with which they were infected, but these antisera were not universally reactive against pneumococcal strains. However, the bacterial isolates and serum samples from these clinical studies provided the critical reagents for the experiments that ultimately led to the identification of DNA as the hereditary material.
Pneumococcal Research Provides Critical Tools in DNA Research
Although numerous scientists engaged in pneumococcal research during the first half of the twentieth century, two of these researchers played an especially important role in the course of events that led to the discovery of DNA as the hereditary material . One of these individuals was Oswald Avery . Avery joined the Rockefeller Institute for Medical Research, now the Rockefeller University, in 1913 as part of a team seeking to develop a therapeutic serum for treating lobular pneumonia. Avery believed that knowledge of the chemical composition of the pneumococcus bacterium was essential for understanding and treating the disease . He perfected his biochemical technique by focusing on the chemical composition of the capsule that surrounded virulent S strains of pneumococci. In his early work, Avery helped establish that polysaccharides were a major component of the pneumococcal capsule and that capsules from different serotypes of pneumococci had distinctive polysaccharide compositions. Avery also concerned himself with the role of capsules in pathogenicity, as capsules were notably absent from the surface of nonvirulent R forms of streptococci. Defying the conventional wisdom of the time, Avery hypothesized that the polysaccharides in the capsules were the actual antigens stimulating the production of antibodies in infected patients (Avery & Goebel, 1933).
Purification of the Active Transforming Principle
When Avery first became aware of Griffith's results, he treated them with skepticism. Other researchers and laboratories, however, were quick to reproduce and build upon Griffith's data. Within a few years, Sia and Dawson (1931) showed that transformation could be carried out in liquid cultures of pneumococci as well as in mice, allowing more precise control of environmental variables in transformation experiments. In 1932, Alloway further demonstrated that the active transforming principle was present in sterile, cell-free extracts prepared from heat-treated pneumococci by filtration. These additional findings convinced Avery that the transforming principle could be identified, and he applied his considerable biochemical expertise to its purification from pneumococcal extracts (Avery et al. , 1944).
A critical aspect of any biochemical purification is the development of an assay, or a way to measure the activity of interest. For their experiments, Avery and his colleagues developed conditions under which R cells could be reliably transformed into S cells using extracts of heat-killed Type III S cells. These same conditions could then be used to measure transforming activity in fractions obtained at different steps in the purification process. To quantify the actual amount of transforming principle in a fraction, each sample was tested at a series of increasing dilutions. These data represent four identical experiments in which Avery and his colleagues tested the ability of the purified factor, designated preparation 44, to transform Type II R cells into Type III S cells. The transforming activity was very concentrated in the extract, since it could be diluted ten-thousand-fold without losing its transforming ability. Fractions that maintained transforming activity at the highest dilutions were deemed to possess the highest concentration of transforming activity. Specifically, when at least 0.01μg of the extract was added to cells, transformation was observed. When any less than 0.01μg was added, the transformation was inconsistent (comparing samples 1 and 3 with 2 and 4) (Figure 2).
Physical Characterization of the Transforming Principle
Avery and his colleagues submitted the purified transforming principle to rigorous physical characterization in order to demonstrate that it possessed the properties expected of DNA (Avery et al. , 1944). The elemental composition of the purified transforming compound was close to the theoretical values for DNA (last row, sodium desoxyribonucleate) (Figure 3). Significantly, the purified principle had a high phosphorous content, which is characteristic of DNA, but not of proteins.
Consistent with these results, the factor gave positive reactions in chemical tests for DNA, but negative or weakly positive reactions in tests for proteins and RNA . Other tests indicated that the transforming principle was a very large molecule that absorbed the same spectrum of ultraviolet light as DNA. However, the most definitive proof that the transforming principle was DNA was its sensitivity to specific enzymes, called DNAses, that specifically degrade different kinds of DNA. Avery and his colleagues were able to show that transforming activity was not destroyed by enzymes that degrade proteins or RNA. At the time, Avery could not obtain samples of pure DNAse. Instead, Avery and his colleagues used crude preparations from animal tissues that were known to contain DNAse activity. They then measured the ability of these various crude preparations to destroy the transforming principle in parallel with measurements of phosphatase, esterase, and DNAse activities in the same extracts. In all cases, the ability of the crude extracts to destroy the transforming principle was proportional to their DNAse activity, measured with pure calf thymus DNA as substrate (Figure 4).
DNA Has the Properties Expected of Genes
References and recommended reading.
- Add Content to Group
Article History
Flag inappropriate.


Email your Friend

- | Lead Editor: Bob Moss

Within this Subject (34)
- Applications in Biotechnology (4)
- Discovery of Genetic Material (4)
- DNA Replication (6)
- Gene Copies (5)
- Jumping Genes (4)
- RNA (7)
- Transcription & Translation (4)
Other Topic Rooms
- Gene Inheritance and Transmission
- Gene Expression and Regulation
- Nucleic Acid Structure and Function
- Chromosomes and Cytogenetics
- Evolutionary Genetics
- Population and Quantitative Genetics
- Genes and Disease
- Genetics and Society
- Cell Origins and Metabolism
- Proteins and Gene Expression
- Subcellular Compartments
- Cell Communication
- Cell Cycle and Cell Division
© 2014 Nature Education
- Press Room |
- Terms of Use |
- Privacy Notice |

Visual Browse
Transformation in Bacteria
Transformation in bacteria was first studied by a scientist Frederick Griffith in 1928 . According to Griffith, the DNA or gene transfer can occur either naturally or artificially from one type of bacteria to another. For example, Transformation of non-virulent strain to a virulent cell or vice versa.
To explain the transformation principle, Griffith performed certain experiments on the mice by taking pathogenic bacteria Streptococcus pneumoniae . Transformation results in gene alteration in the recipient cell, due to the incorporation of free DNA from its surrounding through the cell membrane.
The transformation process is widely used in gene cloning, DNA linkage, generation of cDNA libraries and protein expression. Here, we will discuss the definition, stages, competence in the transformation of bacteria. You will also get to know the transformation principle of the bacteria through the Griffith experiment (transformation experiment).
Content: Transformation in Bacteria
Definition of transformation, streptococcus pneumoniae, strains used in griffith experiment, transformation experiment, transforming factor, stages of transformation.
Transformation can define as the process of taking up of an extracellular or free DNA strand of one bacterial cell ( donor’s cell ) by the competent bacterial cell ( recipient’s cell ). The taking up of the DNA strand occurs either by natural or artificial means. The transformation occurs mostly in the closely related species. Therefore, transformation merely refers to the direct insertion , incorporation and expression of the exogenous DNA in the competent bacterial cell (gets transformed by the inclusion of free DNA).
Classification :
- Kingdom : Bacteria
- Phylum : Firmicutes
- Class : Bacilli
- Order : Lactobacillales
- Family : Streptococcaceae
- Genus : Streptococcus
- Species : pneumoniae

To demonstrate the transformation principle, Frederick Griffith had taken the pathogenic bacteria Streptococcus pneumoniae . Further, he observed two different strains of Streptococcus pneumoniae and named it as S-III and R-II strain.
- S-III strain : It is the smooth strain of Streptococcus pneumonia , which is encapsulated with the polysaccharide. S-III strain will act as virulent or wild strain , as the polysaccharide is a virulent factor.
- R-II strain : It is the rough strain of Streptococcus pneumonia , which lacks the polysaccharide covering. The R-II strain will act as mutant or avirulent strain , as the polysaccharide is absent.
To explain the theory of transformation principle, Frederick Griffith performed a series of experiments where he injected two different strains of Streptococcus pneumoniae into the mice and reported the particular strain’s effect into the mice.
- In his first experiment, Griffith used a rough strain of Streptococcus pneumonia ( R-II ) and injected it into the mice. After doing this, he observed that the R-II strain of bacteria did not affect the mice and the mice lived . Therefore, Griffith named R-II strain as an “ Avirulent strain ”.
- In his second experiment, Griffith used a smooth strain of Streptococcus pneumonia ( S-III ) and injected it into the mice. After doing this, he observed that the S-III strain of bacteria killed the mice. Therefore, Griffith named S-III strain as a “ Virulent strain ”.
- In the third experiment, Griffith used smooth or virulent S-III and subjected it to the heat to destroy the virulence. Then, he injected the heat-killed S-III strain into the mice. After doing this, he observed that the heat-killed S-III strain did not affect the mice and the mice lived . Therefore, Griffith concluded that the virulence of the S-III strain becomes ineffective the heat exposure.
- In the fourth experiment, Griffith used rough R-II strain plus dead or heat-killed S-III strain and injected into the mice. After that, he observed the death of mice. Then, he concluded that the S-III strain had transferred something which transformed the R-II strain into the virulent strain (R-III) and caused the death of mice.

To explain the transformation factor (whether it was a protein or some other component), Avery , Macleod and McCarty performed a series of experiments. Their experiment to identify the transformation of R-II to virulent type can be summarized into the following sequential steps:

- First, they extracted different components like protein, polysaccharide, lipid, RNA and DNA from the heat-killed S-III strain.
- After that, they added R-II strain individually into each test tubes.
- In the third step, they used specific enzymes for the digestion of specific components.
- Then, they injected it to the mice.
After doing this experiment, they observed the death of four mice except for the last one. They concluded that the DNA is the transformation factor which has transformed the R-II strain to the virulent type. Therefore, the DNA is the heritable material that has transferred the virulence from the dead or heat-killed S-III strain to the R-II strain.
As the DNA of S-III or virulent strain is destroyed by the enzyme DNase , there will not be any transformation between the heat-killed S-III strain and the R-II strain, and thus there will be no effect on the mice. They also concluded that even though the polysaccharide is a virulent factor, but still it is not involved in the transformation as it is not heritable.
There are three stages of transformation which include :
- Competence is the first stage where a cell must be competent to take up the DNA. To develop competence, the cell responds to the environmental signal, allowing the binding and penetration of the free DNA .
- The DNA binding is the second stage of transformation in which the exogenous or free DNA binds to the recipient’s cell wall due to developed competence. This stage occurs at the time of incubation of bacterial cell culture on ice. The DNA will bind to the recipient cell wall of bacteria by forming calcium chloride plus a DNA complex.
- DNA integration is the incorporation of the exogenous DNA that has entered to the recipient cell cytoplasm. Therefore, the insertion of foreign DNA into the chromosome of the recipient cell will cause transformation.
To carry out the transformation process, the bacteria should be competent to take up the free DNA. Competence can define as the physiological state , where a recipient cell is in a state where it can respond to the environmental conditions such as starvation and cell density. Therefore, when a cell becomes competent, it can take up the exogenous DNA from the donor’s cell.
In the process of transformation, competence can be of two types :
Natural competence
Artificial competence.
It is a type where a transformation occurs naturally in response to environmental signals and extreme conditions. About 1% of bacteria can develop competence naturally. A set of genes are carried by the naturally competent bacteria. The genes (DNA) then migrate across the cell membrane naturally and infuse within the recipient’s cell.

In this type, a transformation is induced artificially by some chemical or physical methods. Thus, the transformation process is forced or do not occur naturally. Artificial competence can be achieved by both chemical and physical methods. The artificial competence can be achieved by the chemical method through the divalent cation method and physical method through the electroporation.
Divalent cation method : It was first introduced by the two scientists Mandel and Higa in 1970 . In the divalent cation method, the E.coli in the log phase of growth are taken from the culture. Then, E.coli culture is centrifuged. From the E.coli culture, the pellet of bacteria is resuspended in the divalent ion solution like calcium chloride . After that, the culture is kept under cold conditions that result in the weakening of bacteria’s cell surface and allow the binding of free DNA molecule.

Then, the bacterial suspension is suddenly subjected to the high temperature (42 Degrees Celsius) for 30 seconds in the boiling water bath, and the process called heat shock . It results in the thermal imbalance within the bacterial cell and forces the binding of free DNA into the cell.
Electroporation : It is an alternative method of chemical transformation. In electroporation, the bacterial cell is subjected to a high voltage of 15 kV/cm for a 5 µ sec under the influence of an electric field, and the process is called electroshock . The electric shock enhances the ability to take up the free DNA strand. In 1982, a technique of introducing free DNA into the mice was carried out by a scientist Neumann where he treated it with the short pulses at high voltage.

Neumann concluded that the electric shock increases the cell’s membrane potential and thereby increases the cell permeability to take up the charged molecule like DNA.
We can conclude that the non-competent cell has to be competent to carry out the transformation. The competence is developed by the environmental signals like temperature, pH, heat etc., making the cell competent by enhancing the ability to take up the free DNA.
Related Topics:
- Major Groups of Microorganisms
- Bacterial Conjugation
- Nuclear Pore Complex
- Nutrition in Bacteria
Leave a Comment Cancel Reply
Your email address will not be published. Required fields are marked *
Start typing and press enter to search
- Skip to primary navigation
- Skip to main content
- Skip to footer
Biology Wise
Frederick Griffith’s Experiment and the Concept of Transformation
Transformation is a molecular biology mechanism via which foreign and exogenous genetic material is taken up by a cell and incorporated into its own genome. This phenomenon was first described and discovered by British bacteriologist, Frederick Griffith. The concept of transformation and the experiment that led to its discovery are described here.
Like it? Share it!

Did You Know?
Other processes by which exogenous genetic material is taken up by a cell include conjugation (transfer of DNA between two bacterial cells that are in direct contact) and transduction (injection of viral DNA by a bacteriophage into the host bacterial cell).
The post-World War I Spanish influenza pandemic influenced Frederick Griffith to study the epidemiology and pathology of bacterial pneumonia in order to attempt creating a successful vaccine. Hence, he carried out experiments, where he injected mice with strains of virulent and avirulent Streptococcus pneumoniae. The experiment he reported in 1928, gave the first description of the phenomenon of transformation, where one bacterial strain could change into the other strain, and this activity was linked to an unidentified element called the transforming factor or transforming principle.
Oswald T. Avery, an American pneumococcal researcher, speculated that Griffith’s experiment lacked appropriate control. However, subsequent, similar experiments carried out in Avery’s laboratory confirmed Griffith’s discovery. The experiments conducted later by Avery, MacLeod, and McCarty, and by Hershey and Chase proved that the transforming factor was DNA and elucidated its exact nature. Thereby, establishing the central role of DNA in inheritance.
Frederick Griffith’s Experiment

For his experiments, Griffith used two strains of Streptococcus pneumoniae that affected mice – type III S (smooth) and type II R (rough). The type III S form has a smooth appearance due to the presence of a polysaccharide layering over the peptidoglycan cell wall of the bacterial cell. This extra coating helps the cell in evading the phagocytosis carried out by the immune cells of the host; hence, allowing the strain to proliferate and become virulent.
In contrast, the type II R form lacks this coating, and hence, has a rough appearance. The absence of the polysaccharide layer leads to its efficient elimination by the host’s immune cells rendering the strain avirulent. While injecting the mice with these bacteria, Griffith devised four sets of inoculation that are as follows:
Type III S bacteria When the mice were inoculated, the bacterial virulence was exhibited, causing pneumonia, and this eventually led to the death of the mice. On examining the blood of the deceased mice, progeny of the inoculated cells were obtained.
Type II R bacteria When injected into the mice, the bacterial cells were successfully eliminated by the immune system, and hence, the mice lived. The blood showed no presence of the inoculated cells.
Type III S heat-killed bacteria When the virulent strain was rendered avirulent by heating and killing it (heat-killed), and then injected into the mice, the strain did not show virulence, and was eliminated by the host’s immune system; hence, the mice survived. Their blood showed no presence of the inoculated cells.
Type II R bacteria + Type III S heat-killed bacteria Injecting the mice with a combination of equal number of cells of type II R strain and heat-killed type III S strain, caused pneumonia which progressed till the mice died. The bacterial cells isolated from the blood of these mice showed the presence of live type III S bacterial cells.
This indicated that the live R strain had assimilated and incorporated the virulent element from the heat-killed S strain in order to transform itself into the virulent S strain. Based on this observation, Griffith concluded that a transforming element from the heat-killed strain was accountable for the transformation of the avirulent strain into the virulent strain. Successive experiments carried out in 1944 by Oswald Avery, Colin MacLeod, and Maclyn McCarty, proved that the element taken up by the harmless strain was genetic in nature.
Concept of Transformation
Transformation is a stable genetic change brought about by the uptake of naked DNA, and the state of being able to take up exogenous DNA is called competence. They occur in two forms―natural and artificial.
Natural Transformation
Only 1% of the bacterial species is capable of taking up DNA. Bacteria also exchange genetic material through a process called horizontal gene transfer, where bacterial cells conjugate and form a bridge via which the genetic material is transferred from one cell to another. A few bacterial species also release their DNA via exocytosis on their death, and this DNA is later taken up by the bacterial cells present in the vicinity. While transformation can occur between various bacterial species, it is most efficient when occurring between closely related species. These cells possess specific genes that code for natural competence allowing them to transport the DNA across the cellular membrane and into the cell. This transport involves the proteins associated with the type IV pili and type II secretion system as well as the DNA translocase complex at the cytoplasmic membrane.
This mechanism differs slightly due to the difference in the structure of the cell membranes of the bacteria. Bacteria are broadly classified into two types based on this difference – Gram-negative and Gram-positive. The general outline is more or less similar. The presence of exogenous DNA is detected by the cell and natural competence is induced, then the foreign DNA binds to a DNA receptor on the surface of these competent cells. This receptor binding allows the activation of the DNA translocase system that allows the passing of DNA into the cell via the cell membrane. During this process, one strand of the DNA is degraded by the action of nucleases. The translocated single strand is then incorporated into the bacterial genome via the help of a RecA-dependent process.
Gram-negative bacteria show the presence of an extra membrane, hence, for DNA to be taken up, a channel is formed on the outer membrane by secretins. The uptake of a DNA fragment is generally not specific to its sequence; however, in some bacterial species, it has been seen that the presence of certain DNA sequences facilitate and enhance efficient uptake of the genetic material.
Artificial Transformation
It is carried out in laboratories in order to carry out gene expression studies. To impart competence, the cells are incubated in a solution containing divalent cations (calcium chloride) under cold conditions, and then, exposed to intermittent pulses of heat. The concentration of the solution depends on the protein and liposaccharide content of the membrane, and the intensity of the heat pulses varies according to the time duration of the pulses, i.e., high intensity pulses should be for very short periods; whereas, low intensity pulses can be for longer durations.
The divalent cations function to weaken the molecular structure of the cell membrane, hence, making it more permeable. The subsequent heat pulses cause the creation of a thermal imbalance, and in the process of regaining balance, the DNA molecules gain entry via the weakened membrane and into the cell.
Artificial competence can be alternatively induced and promoted via the use of a technique called electroporation. It involves applying an electric current to the cell suspension. This causes the formation of pores in the cell membrane. The exogenous DNA is taken up via these holes, which are resealed via the cell membrane repair machinery.

Saccharomyces cerevisiae can be transformed by exogenous DNA using various methods. Yeast cells are treated with certain digesting enzymes that degrade the cell walls. This yields naked cells (devoid of cell wall) called spheroplasts. They are extremely fragile but have a high frequency of foreign DNA uptake.
Another method that can be used is, exposing the cells to alkaline cations such as lithium (from lithium acetate) and PEG. The PEG helps in pore formation, and the cations in the transport of the DNA fragment inside the cell.
The process of electroporation can also be used for transformation purposes, and efficiency can be enhanced using enzymatic digestion or agitation using glass beads.
The most common method of transforming plant cells is the Agrobacterium mediated transfer. In this method, the tissue of cells to be transformed is cut up into small uniform pieces, and then, treated with a suspension containing Agrobacterium. The foreign DNA gains entry via the cuts on the tissue, and the wound healing compounds secreted from the cuts, activate the virulence operon of the Agrobacterium. his causes the Agrobacterium. to infect the tissue and carry out its normal action of tumor induction. This function allows the transformed plant cells to proliferate. The cells are grown on a selective media till the transformed cells grow into plantlets with shoots and roots. They are then planted in soil and allowed to grow naturally.
Plant cells can also be transformed using viral particles (transduction). Here, the genetic material to be inserted is packaged into a suitable plant virus. This modified virus is then allowed to infect the plant cells. The transfer occurs according to the viral machinery and transformation is achieved. Electroporation can also be used for plant cells.
Introduction of foreign DNA into animal cells is conducted using viral or chemical agents like the ones used in case of plant and bacterial cells. However, since the term transformation is also used to refer to the progression of cancerous growth in animals, here, the term transfection is used.
This concept and technique has seen varied applications in the field of molecular biology with respect to expression studies, gene knockout studies, and cloning experiments. It is also used in the production of genetically modified organisms.
Get Updates Right to Your Inbox
Privacy overview.
Scan to visit
- Introduction to Genomics
- Educational Resources
- Policy Issues in Genomics
- The Human Genome Project
- Funding Opportunities
- Funded Programs & Projects
- Division and Program Directors
- Scientific Program Analysts
- Contacts by Research Area
- News & Events
- Research Areas
- Research Investigators
- Research Projects
- Clinical Research
- Data Tools & Resources
- Genomics & Medicine
- Family Health History
- For Patients & Families
- For Health Professionals
- Jobs at NHGRI
- Training at NHGRI
- Funding for Research Training
- Professional Development Programs
- NHGRI Culture
- Social Media
- Broadcast Media
- Image Gallery
- Press Resources
- Organization
- NHGRI Director
- Mission and Vision
- Policies and Guidance
- Institute Advisors
- Strategic Vision
- Leadership Initiatives
- Diversity, Equity, and Inclusion
- Partner with NHGRI
- Staff Search
1944: DNA is \"Transforming Principle\"
1944: dna is "transforming principle".
Avery, MacLeod and McCarty identified DNA as the "transforming principle" while studying Streptococcus pneumoniae , bacteria that can cause pneumonia. The bacteriologists were interested in the difference between two strains of Streptococci that Frederick Griffith had identified in 1923: one, the S (smooth) strain, has a polysaccharide coat and produces smooth, shiny colonies on a lab plate; the other, the R (rough) strain, lacks the coat and produces colonies that look rough and irregular. The relatively harmless R strain lacks an enzyme needed to make the capsule found in the virulent S strain.
Griffith had discovered that he could convert the R strain into the virulent S strain. After he injected mice with R strain cells and, simultaneously, with heat-killed cells of the S strain, the mice developed pneumonia and died. In their blood, Griffith found live bacteria of the deadly S type. The S strain extract somehow had "transformed" the R strain bacteria to S form. Avery and members of his lab studied transformation in fits and starts over the next 15 years. In the early 1940s, they began a concerted effort to purify the "transforming principle" and understand its chemical nature.
Bacteriologists suspected the transforming factor was some kind of protein. The transforming principle could be precipitated with alcohol, which showed that it was not a carbohydrate like the polysaccharide coat itself. But Avery and McCarty observed that proteases - enzymes that degrade proteins - did not destroy the transforming principle. Neither did lipases - enzymes that digest lipids. They found that the transforming substance was rich in nucleic acids, but ribonuclease, which digests RNA, did not inactivate the substance. They also found that the transforming principle had a high molecular weight. They had isolated DNA. This was the agent that could produce an enduring, heritable change in an organism.
Until then, biochemists had assumed that deoxyribonucleic acid was a relatively unimportant, structural chemical in chromosomes and that proteins, with their greater chemical complexity, transmitted genetic traits.
« Previous Event | Next Event »
Last updated: April 23, 2013
IMAGES
VIDEO
COMMENTS
Frederick Griffith experiments were conducted with Streptococcus pneumoniae. During the experiment, Griffith cultured Streptococcus pneumoniae bacteria which showed two patterns of growth. One culture plate consisted of smooth shiny colonies (S) while other consisted of rough colonies (R).
Griffith's experiment, [1] performed by Frederick Griffith and reported in 1928, [2] was the first experiment suggesting that bacteria are capable of transferring genetic information through a process known as transformation.
British bacteriologist Frederick Griffith reported the first demonstration of bacterial transformation—a process in which external DNA is taken up by a cell, thereby changing its morphology and physiology.
Frederick Griffith Discovers Bacterial Transformation. Figure 1: R variant phenotypes. In this figure the colonies of R variant on the left are grown on agar in the absence of the...
The "Griffith's Experiment," carried out by English bacteriologist Frederick Griffith in 1928, described the transformation of a non-pathogenic pneumococcal bacteria into a virulent strain. Griffith combined living non-virulent bacteria with a heat-inactivated virulent form in this experiment.
Frederick Griffith carried out his experiment with mice and Streptococcus pneumoniae (pneumococcus bacteria), the causative agent of pneumonia in humans. And he showed that there was a transforming principle in bacterial genome.
The scientist Frederick Griffith observed that a living non-virulent bacterial strain could be transformed into a virulent strain after it was mixed with a dead virulent strain. © 2013 Nature...
Transformation in bacteria was first studied by a scientist Frederick Griffith in 1928. According to Griffith, the DNA or gene transfer can occur either naturally or artificially from one type of bacteria to another.
Transformation is a molecular biology mechanism via which foreign and exogenous genetic material is taken up by a cell and incorporated into its own genome. This phenomenon was first described and discovered by British bacteriologist, Frederick Griffith.
Griffith had discovered that he could convert the R strain into the virulent S strain. After he injected mice with R strain cells and, simultaneously, with heat-killed cells of the S strain, the mice developed pneumonia and died. In their blood, Griffith found live bacteria of the deadly S type.